Also see an excellent Scholarpedia article here
Systems and Behavioural Neuroscience Laboratory
- The Otolith Organs
- The Semi-Circular Canals
- The Functions of the Vestibular System
- The Vestibulo-Ocular Reflex
 |
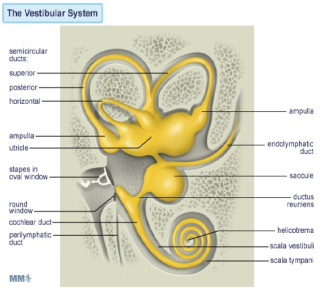
Introduction to the Vestibular System The vestibular system is phylogenetically the oldest part of the inner ear. It is situated in the petrous part of the temporal bone, in close proximity to the cochlea. The vestibular system responds to movement of the head relative to space and gravity, using inertial-sensing receptors which are activated by forces arising from the acceleration of mass in accordance with Newton’s law: Force = Mass X Acceleration In order to determine the absolute movement of a body in three dimensional space, reliable information is required about movement in each of the 6 "degrees of freedom" permitted in three dimensional space, i.e. three translation or straight lines (up-down, left-right, fore-aft), and three rotational (in one horizontal, and two vertical planes at right angles to each other) movements. There is one vestibular system on each side of the head, in close approximation to the cochlea. |
|
Each side of this bilateral system consists of two types of sensors:
The utricle and the saccule are two sac-like structures each of which contains a specialized region (the macula) which is made up of a ciliated sensory epithelium (the vestibular hair cells). In humans, the hair cells in the vestibular system differ somewhat from those in the auditory system, in that each vestibular cell, in addition to having a number of thin stereo-cilia, also has one thicker longer kino-cilium positioned at one end of the cell's hair-bearing surface. |
 |
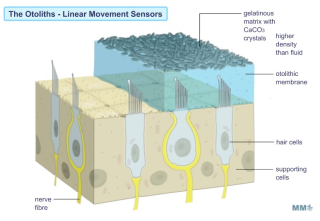 |
The hair cells of the vestibular system also exhibit a constant "resting discharge activity" even in the absence of a stimulus. Thus, stimulation is sensed by the central nervous system as a change in this resting, "spontaneous" discharge rate. The cilia which emerge from the hair cells are embedded in a gelatinous matrix containing solid CaCO3 crystals (the otoconia) which overlies the cells. During linear acceleration, the crystals (being denser than the surrounding fluid) will tend to be left behind due to their inertia. It has been demonstrated that the resultant bending of the cilia causes cell excitation when the bending is toward the kino-cilium (with a resultant increase in the firing frequency of the innervated afferent sensory fibres of the VIII th nerve), and inhibition when away from the kino-cilium (with a resultant decrease in the firing frequency of the innervated afferent sensory fibres of the VIII th nerve). |
Since they are sensitive to acceleration, the otolith organs detect the direction and magnitude of gravity, as well as transient linear accelerations due to movement (for example: tilting the head produces a transient linear acceleration which is reflected in changes in the firing frequency of afferent fibres.
innervating the sensory cells).
The Semi-Circular Canals
The three semicircular canals are more or less orthogonal (at right angles) with respect to each other and are continuous with the utricle and the saccule. The horizontal canals are approximately horizontal when the head is kept at its normal attitude. The vertical canals are arranged in diagonal planes which subtend roughly 45 degrees relative to the sagittal and frontal planes of the skull. Thus the anterior canal on one side of the head is parallel to the posterior canal on the other. Each canal is comprised of a circular path of fluid continuity, interrupted at the ampulla (where the sensory receptor cells are located in the crista) by a water tight, swing-door-like elastic membrane called the cupula. The ampulla contains the sensory receptor cells of the semicircular canals: the vestibular hair cells.
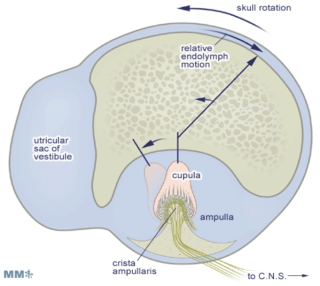
Rotational angular acceleration of the whole canal causes fluid to be left behind on account of its inertia; which, in turn, deflects the cupula. As in the otoliths, the resultant bending of the cilia causes cell excitation when the bending is toward the kino-cilium (with a resultant increase in the firing frequency of of the VIII th nerve fibres), and inhibition when away from the kino-cilium (with a resultant decrease in the firing frequency of the VIII th nerve fibres).
This process is similar to the transduction process already described for the cochlea and otolith organs above. The sensory epithelium of the crista is very similar to that found in the otolith organs, with the exception that the cilia are much longer, extending far up into the cupula.
Again, as with the otolith organs, the sensory cells exhibit a "resting discharge" that is modified (increased or decreased) depending on the direction in which the cupula is deflected (i.e. depending on the direction of rotation). In contrast to the otolith organs, the cilia-kinocilium complexes of the hair cells in the semicircular canals are oriented in one direction, such that all the hair cells in each canal one of the three canals are maximally excited by the same direction of angular rotation.
Although the stimulus on the semicircular canals is angular (rotational) acceleration, the neural output from the sensory cells in the ampulla represents the velocity at which the canal is being rotated over the range of normal head movements. The canal mechanism therefore performs a mathematical integration of the input signal (i.e. the integral of acceleration = velocity). This comes about largely due to the very small size of the canal (the thin tube has a diameter of about 0.3 mm in humans), which results in a large increase in the viscous properties of the fluid.
Thus, the semicircular canal system acts as a precise angular (rotational) speedometer: its neural output is directly proportional to the angular (rotational) velocity of head movements. By combining the input from each of the three canals, the brain can create a representation of the vector which describes the instantaneous speed of head rotation relative to 3-dimensional space.
The Functions of the Vestibular System
The information from the vestibular apparatus is used in three ways:
To provide a subjective sensation of movement and/or displacement in 3-dimensional space.
For example, the hair cells of the utricle provide a sensation of head tilt based on the direction in which the cilia are bent by the gravitational force. When the head is tilted in the direction of polarity of a given cell, it depolarizes and excites the afferent fiber. Alternatively, when the head is tilted in the opposite direction, the same cell hyperpolarizes and inhibits the afferent fiber.
To maintain upright body posture (balance).
A variety of reflexes of the limb musculature, are mediated by activation of the otolith organs and semicircular canals. When the vestibular system is activated these reflexes result in the stabilization of the head's position in space (vestibulospinal and otolith-spinal reflexes).
To control the muscles that move the eyes, so that in spite of the changes in head position which occur during normal activities such as walking and running, the eyes remain stabilized on a point in space . The eye movements which are generated by activation of the vestibular system are called vestibulo-ocular reflexes and are discussed in greater detail on the next page.
The Vestibulo-Ocular Reflex
Unblurred vision is only possible if the eye is stationary (fixed) with respect to a viewed object. The vestibulo-ocular reflex (VOR) is an important mechanism by which unblurred vision is made possible during head movements that are generated during everyday activities such as walking and running. For example, if the head is turned to the left, this reflex causes the eyes to move to the right (i.e. in the opposite direction of the head movement). The oppositely directed eye movement occurs at the same velocity as the head movement, and therefore generates an eye movement which keeps our line of sight fixed on the same point in visual space both during and following the movement.
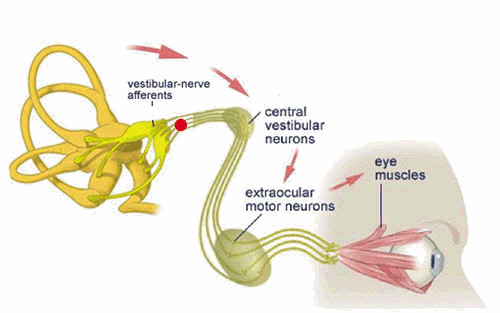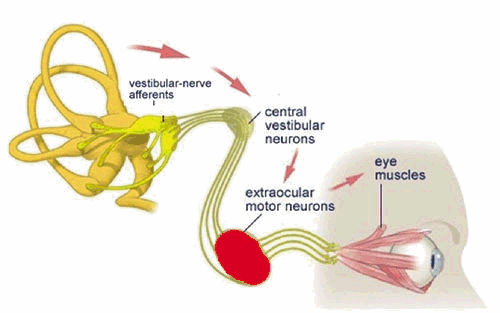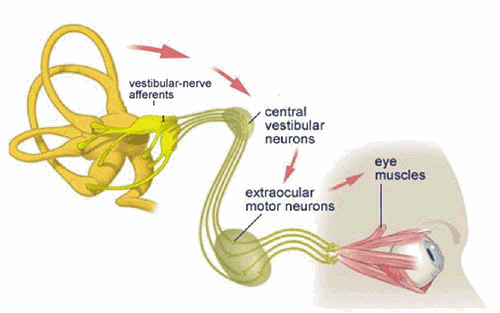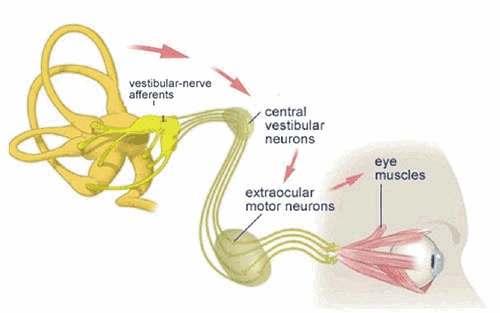
During short head movements, these compensatory eye movements remain well within the mechanical limits of eye rotation. However during large amplitude head rotation, the eye can reach its limit of excursion long before the head movement is completed. Consequently, during this condition, an additional feature is added to the VOR: when the eye reaches an extreme position, it is rapidly flicked back to a new starting position. From this new starting position, the eye then continues a new cycle of compensatory movement during continuing head movement. The resulting "saw tooth" pattern of slow compensatory/ rapid resetting eye movements (slow phases and quick phases respectively) are referred to as vestibular nystagmus.